The emergence of eukaryotic cells was important in the evolution of complex multicellular life. But how did eukaryotes evolve?
By KAREN S KABNICK and DEBRA A PEATTIE
Today both eukaryotes and prokaryotes still exist. Eukaryotes can be found variously as single-celled organisms called protists, and as organized systems in multicellular organisms. The cells of all plants, animals and fungi are eukaryotes. The prokaryotes live on in the two major divisions of bacteria-the eubacteria and the archebacteria. As in earlier times, prokaryotes are less likely than eukaryotes to form organisms larger than a single cell.
Not only do eukaryotic cells allow larger and more complex organisms to be made, but they are themselves larger and more complex than prokaryotic cells. Whether eukaryotic cells live singly or as part of a multicellular organism, their activities can be much more complex and diversified than those of their prokaryotic counterparts. In prokaryotes, all internal cellular events take place within a single compartment, the cytoplasm. Eukaryotes contain many subcellular compartments called organelles. Even single-celled eukaryotes can display remarkable complexity of function; some have features as specialized and diverse as sensory bristles, mouth parts, muscle-like contractile bundles, or stinging darts.
 Much evidence from the fossil record and from molecular biology indicates that eukaryotes evolved from
prokaryotes. But the details of this important transition are difficult to trace, having happened so long ago that most evidence of it has disappeared. One way we can devise plausible scenarios for the evolution of eukaryotes is to explore the vestiges of older systems that remain in existing cells and reconstruct how things might have happened. Especially useful for this purpose are many of the single-celled eukaryotes whose life-styles in
Much evidence from the fossil record and from molecular biology indicates that eukaryotes evolved from
prokaryotes. But the details of this important transition are difficult to trace, having happened so long ago that most evidence of it has disappeared. One way we can devise plausible scenarios for the evolution of eukaryotes is to explore the vestiges of older systems that remain in existing cells and reconstruct how things might have happened. Especially useful for this purpose are many of the single-celled eukaryotes whose life-styles in  some ways resemble those of their prokaryotic forebears, and which are thought to be among the earliest eukaryotes that are still in existence. Studies of such organisms have led to two main theories, which are not mutually exclusive, that seek to explain how eukaryotes evolved. Recent evidence suggests that one single-celled organism, the intestinal parasite Giardia lamblia, represents the first line of descent from the ancestral cells that took on eukaryotic features. As such, a study of the cell biology of this "missing link" organism can help to answer many questions about the way things might have been early in the history of life.
some ways resemble those of their prokaryotic forebears, and which are thought to be among the earliest eukaryotes that are still in existence. Studies of such organisms have led to two main theories, which are not mutually exclusive, that seek to explain how eukaryotes evolved. Recent evidence suggests that one single-celled organism, the intestinal parasite Giardia lamblia, represents the first line of descent from the ancestral cells that took on eukaryotic features. As such, a study of the cell biology of this "missing link" organism can help to answer many questions about the way things might have been early in the history of life.
Prokaryotes and Eukaryotes
On a very fundamental level, eukaryotes and prokaryotes are similar. They share many aspects of their basic chemistry, physiology and metabolism. Both cell types are constructed of and use similar kinds of molecules and macromolecules to accomplish their cellular work. In both, for example, membranes are constructed mainly of fatty substances called lipids, and molecules that perform the cell's biological and mechanical work are called proteins.
Eukaryotes and prokaryotes both use the same chemical relay system to make protein. A permanent record of the code for all of the proteins the cell will require is stored in the form of DNA. Because DNA is the master copy of the cell's (or organism's) genetic make-up, the information it contains is absolutely crucial to the maintenance and perpetuation of the cell. As if to safeguard this archive, the cell does not use the DNA directly in protein synthesis but instead copies the information onto a temporary template of RNA, a chemical relative of DNA. Both the DNA and the RNA constitute a "recipe" for the cell's proteins. The recipe specifies the order in which amino acids, the chemical subunits of proteins, should be strung together to make the functional protein. Protein synthesis both in eukaryotes and prokaryotes takes place on structures called ribosomes, which are composed of RNA and protein. This illustrates one way in which prokaryotes and eukaryotes are similar and highlights the idea that differences between these organisms are often architectural. In other words, both cell types use the same bricks and mortar, but the structures they build with these materials vary dramatically.
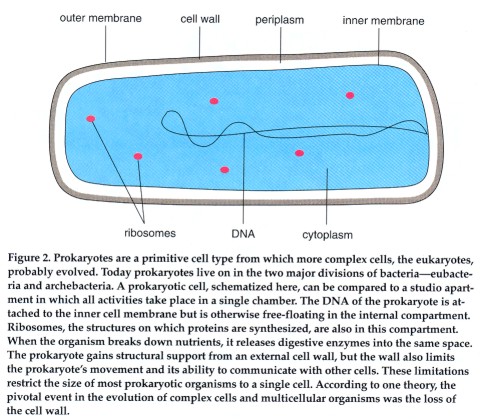 The prokaryotic cell can be compared to a studio apartment: a one-room living space that has a kitchen area abutting the living room, which converts into a bedroom at night. All necessary items fit into their own locations in the one room. There is an everyday; washable rug. Room temperature is comfortable-not too hot, not too cold. Conditions are adequate for everything that must occur in the apartment, but not
optimal for any specific activity. In a similar way, all of the prokaryote's functions fit into a single compartment. The DNA is attached to the cell's membrane. Ribosomes float freely in the single compartment. Cellular respiration-the process by which nutrients are metabolized to release energy-is carried out at the cell membrane; there is no dedicated compartment for respiration.
The prokaryotic cell can be compared to a studio apartment: a one-room living space that has a kitchen area abutting the living room, which converts into a bedroom at night. All necessary items fit into their own locations in the one room. There is an everyday; washable rug. Room temperature is comfortable-not too hot, not too cold. Conditions are adequate for everything that must occur in the apartment, but not
optimal for any specific activity. In a similar way, all of the prokaryote's functions fit into a single compartment. The DNA is attached to the cell's membrane. Ribosomes float freely in the single compartment. Cellular respiration-the process by which nutrients are metabolized to release energy-is carried out at the cell membrane; there is no dedicated compartment for respiration.
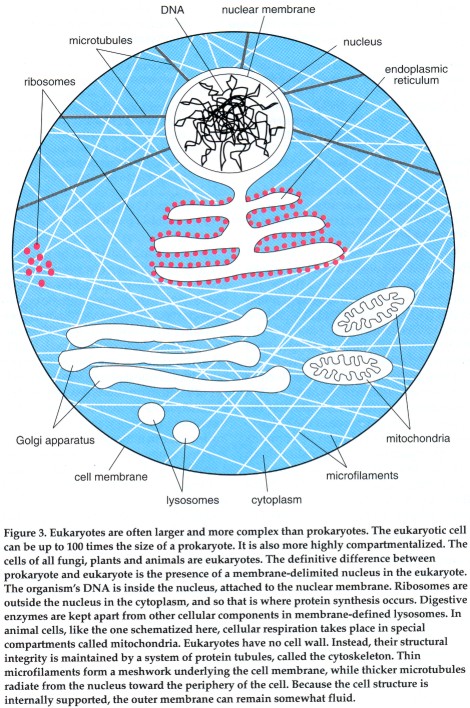 A eukaryotic cell can be compared to a mansion, where specific rooms are designed for particular activities. The mansion is more diverse in the activities it supports than the studio apartment. It can accommodate overnight guests comfortably and support social activities for adults in the living room or dining room, for children in the playroom. The baby's room is warm and furnished with bright colors and a soft, thick carpet. The kitchen has a stove, a refrigerator and a tile floor. Items are kept in the room that is most appropriate for them, under conditions ideal for the activities in that specific room.
A eukaryotic cell can be compared to a mansion, where specific rooms are designed for particular activities. The mansion is more diverse in the activities it supports than the studio apartment. It can accommodate overnight guests comfortably and support social activities for adults in the living room or dining room, for children in the playroom. The baby's room is warm and furnished with bright colors and a soft, thick carpet. The kitchen has a stove, a refrigerator and a tile floor. Items are kept in the room that is most appropriate for them, under conditions ideal for the activities in that specific room.
A eukaryotic cell resembles a mansion in that it is subdivided into many compartments. Each compartment is furnished with items and conditions suitable for a specific function, yet the compartments work together to allow the cell to maintain itself, to replicate and to perform more specialized activities.
Taking a closer look, we find three main structural aspects that differentiate prokaryotes from eukaryotes. The definitive difference is the presence of a true (eu) nucleus (karyon) in the eukaryotic cell. The nucleus, a double-membrane casing, sequesters the DNA in its own compartment and keeps it separate from the rest of the cell. In contrast, no such housing is provided for the DNA of a prokaryote. Instead the genetic material is tethered to the cell membrane and is otherwise allowed to float freely in the cell's interior. It is interesting to note that the DNA of eukaryotes is attached to the nuclear membrane, in a manner reminiscent of the attachment of prokaryotic DNA to the cell's outer membrane.
Although DNA performs the same critical function in both cell types, the presence or absence of a nucleus has some profound implications for the form that the molecule takes and the way that the DNA template ultimately becomes translated into protein. In prokaryotes almost all of the organism's genetic information is carried on a single circular piece of DNA. The genetic material of the eukaryotic cell, on the other hand, consists of several 1inear pieces of DNA. The exact number of linear DNA segments varies from species to species. Generally, the DNA in a eukaryotic cell looks like a loose tangle of yarn, except during cell division, when the DNA becomes tightly wrapped into the structures called chromosomes. The membrane surrounding the eukaryotic cell's nucleus breaks apart during cell division and reappears intact in the daughter cells, one nucleus in each daughter.
Not only is the physical configuration of the DNA different in the two cell types, but they also differ in the number of sets of genetic instructions they contain. A prokaryotic cell contains only a single representation of the genetic information the organism requires; in this condition the cell is said to be haploid. In contrast, most eukaryotes have two sets of genetic information during some stage of their lives. Cells containing two sets of genetic information are referred to as diploid. Some simple eukaryotes pass through only a fleeting diploid stage, but higher eukaryotes spend most of their lives as diploid cells. Multicellular organisms can include both diploid and haploid eukaryotic cells. Most of the cells of the organism's body are diploid. The gametes--eggs and sperm-are haploid eukaryotes. During fertilization two haploid gametes fuse, thus restoring the diploid condition to the resulting embryo. Having two sets of genetic information offers the eukaryote certain advantages over prokaryotes, and the emergence of the diploid state was an important milestone in evolution.
The nucleus is one of several specialized compartments in the eukaryotic cell. Other compartments, called organelles, accommodate several other cellular activities. Prokaryotes do not have subcellular compartments, and this constitutes the second major distinction between the two cell types.
The organelles of eukaryotes include membrane-bounded compartments such as the lysosome, a highly acidic compartment in which digestive enzymes break down food. The endoplasmic reticulum is an interconnected system of membranes in which lipids are synthesized and some proteins are chemically modified. The endoplasmic reticulum communicates with another membrane system called the Golgi apparatus, where proteins are further processed and marked for transport to various sites inside or outside the cell. Eukaryotic cells contain special energy centers. In animal cells these are the mitochondria; plant cells have chloroplasts as well as mitochondria. Within mitochondria, organic compounds are broken down to generate the energy-rich molecule adenosine triphosphate (ATP). ATP is a sort of molecular fuel, which when degraded provides energy for many of the cell's biochemical reactions. ATP is also generated in the chloroplasts of plant cells, but the energy for its synthesis is derived from sunlight, in a process that also builds up carbohydrates and liberates oxygen.
The third distinguishing feature between the two cell types is the way in which the cell maintains its shape. Cells, like most animals, have skeletons. And, as in many animals, the cellular skeleton can be either internal or external. Prokaryotes have an external skeleton; a strong wall of cross-linked sugar and protein molecules surrounds the cell membrane and is made rigid by the turgor pressure of the cell. The wall lends structural support. It is also impermeable to many macromolecules and thus helps to maintain a barrier between substances inside and outside the cell. Such an external skeleton limits the ability of the prokaryotic cell to move. It also limits communication between cells, a condition that probably accounts for the vastly decreased ability of prokaryotes to form multicellular organisms.
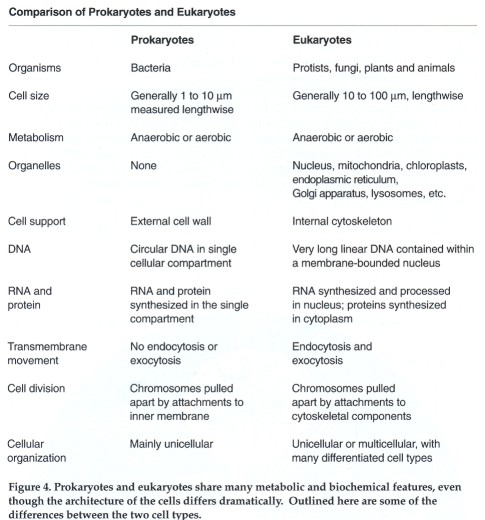 The skeleton of the eukaryotic cell is internal; it is formed by a complex of protein tubules called the cytoskeleton. The internal placement of the cytoskeleton means the surface exposed to the environment is a pliable membrane rather than a rigid cell wall. The combination of an internal framework and a nonrigid outer membrane expands the repertory of motion and activity of the eukaryotic cell. For example, the cell can contract, as does a muscle cell. (The cells of most higher plants have a wall even more rigid than the prokaryotic wall. Plant cell walls are chemically and structurally very different from prokaryotic walls; presumably they are a later, independent adaptation.)
The skeleton of the eukaryotic cell is internal; it is formed by a complex of protein tubules called the cytoskeleton. The internal placement of the cytoskeleton means the surface exposed to the environment is a pliable membrane rather than a rigid cell wall. The combination of an internal framework and a nonrigid outer membrane expands the repertory of motion and activity of the eukaryotic cell. For example, the cell can contract, as does a muscle cell. (The cells of most higher plants have a wall even more rigid than the prokaryotic wall. Plant cell walls are chemically and structurally very different from prokaryotic walls; presumably they are a later, independent adaptation.)
The Way It Was
Two billion years ago, before the emergence of the first eukaryotes, life on earth was very different from what it is today. The organisms populating the earth were prokaryotes, similar to modem bacteria. But unlike the vast majority of modem organisms, even modem prokaryotes, these primordial organisms did not use oxygen. Free oxygen was scarce on the primordial earth, and the earliest organisms evolved a metabolism based on sulfur and hydrogen sulfide (H2S) rather than oxygen and water (H2O). Many of these organisms were obligate anaerobes: Not only did they fail to make use of oxygen, but they could not survive in its presence.
How did eukaryotic cells, as well as modem aerobic prokaryotes, evolve from these anaerobic forerunners? Because of the importance of the eukaryotic cell in the evolution of complex living organisms, the question is of intense interest. Two theories offer competing explanations. Although the theories differ dramatically, they are not mutually exclusive. We can envisage a scenario in which both mechanisms contribute to the evolution of the eukaryotes.
You Are What You Eat
The first theory; proposed by Lynn Margulis of the University of Massachusetts at Amherst and others, suggests that a dramatic increase in atmospheric oxygen and the transition from anaerobe to aerobe were of primary importance in driving the evolution of eukaryotes. According to this theory, a population of primitive bacteria acquired the ability to photosynthesize. Photosynthetic organisms can use the energy of light to convert carbon dioxide into sugar. The sugar is then used as fuel to make ATP, which in turn provides energy for many other biochemical processes. A byproduct of photosynthesis is oxygen, and so a consequence of the evolution of photosynthetic bacteria was an overwhelming increase in atmospheric oxygen. The increasing abundance of oxygen posed a problem for those creatures that could not make metabolic use of it. Oxygen was toxic to these obligate anaerobes, and their continued survival depended on their developing the ability to use it, or at least to tolerate it.
Organisms that adapted to oxygen did so relatively quickly, according to Margulis's theory. Margulis proposes that an anaerobic organism engulfed a smaller aerobic organism believed to be the ancestor of mitochondria. The ingested aerobes were not digested, but lived on inside their new host. Hence was born a mutually beneficial arrangement, where the small aerobe would convert oxygen into ATP for the energetic benefit of both cells. The host thus became tolerant to oxygen. In turn, the host organism eventually performed other functions, such as protein synthesis, for the aerobe.
The vestiges of that early union appear to live on today in modem eukaryotes. The mitochondria of both plant and animal cells and the chloroplasts of plants have many features that suggest they may have descended from free-living bacteria-like organisms. Both organelles have their own DNA, and both organelles replicate independently of cell division.
![]() The theory advanced by Margulis postulates that other eukaryotic organelles were similarly derived from free-living organisms that became symbiotic with larger organisms. For example, the flagellum that helps to propel some eukaryotes by its whipping motion may have evolved from a primitive, unicellular, spiral-shaped organism, like a spirochete, which became associated with a larger host. The term "serial endosymbiosis" is used to describe the successive incorporation of smaller cells into a larger host to the benefit of both organisms.
The theory advanced by Margulis postulates that other eukaryotic organelles were similarly derived from free-living organisms that became symbiotic with larger organisms. For example, the flagellum that helps to propel some eukaryotes by its whipping motion may have evolved from a primitive, unicellular, spiral-shaped organism, like a spirochete, which became associated with a larger host. The term "serial endosymbiosis" is used to describe the successive incorporation of smaller cells into a larger host to the benefit of both organisms.
Two major conclusions follow from the hypothesis of endosymbiotic evolution. One consequence is that the transition from prokaryote to eukaryote would have been relatively quick on the evolutionary time scale. Furthermore, if eukaryotic evolution occurred by endosymbiosis, then the first eukaryotes must have been aerobic. Anaerobic eukaryotes do exist, even today, but the endosymbiotic theory would hold that aerobic eukaryotes evolved first; some of these lost their mitochondria as a result of further selection pressures, giving rise later to anaerobic eukaryotes. Both of these points have generated some controversy, and we shall return to them later.
One important aspect of eukaryotic cells that is not addressed by an endosymbiotic mechanism is the origin of the cell nucleus and other membrane-defined systems such as the endoplasmic reticulum and the Golgi apparatus. Thomas Cavalier-Smith of the University of British Columbia in Vancouver has proposed a second theory that seeks to address the origins of membrane-delimited organelles as well as anaerobic and aerobic eukaryotes.
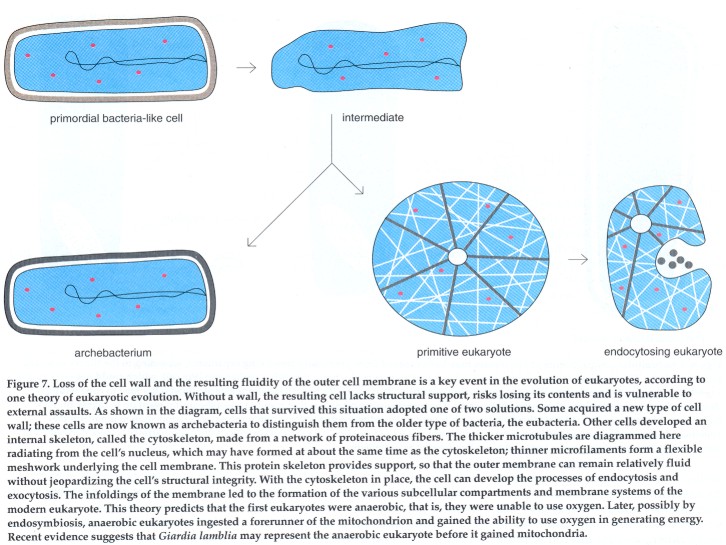 Cavalier-Smith suggests that early prokaryotes were set en route to becoming eukaryotes when they lost the ability to manufacture muramic acid, an essential component of the cell wall in most bacteria. The loss of this sugar causes the rigid cell wall to break down, leaving the cell without support. Cells in this situation are vulnerable to external assaults and risk spilling their contents into the extra- cellular space. The organisms evolved two strategies for coping with the loss of muramic acid and thus diverged into two subsequent lines of descent. A new group of prokaryotes, now called archebacteria, developed a new type of rigid cell wall built without muramic acid. Cells that did not develop a new type of wall evolved an internal proteinaceous skeleton; that is, a cytoskeleton. It is at this point, according to Cavalier-Smith, that the modem eukaryote got its start.
Cavalier-Smith suggests that early prokaryotes were set en route to becoming eukaryotes when they lost the ability to manufacture muramic acid, an essential component of the cell wall in most bacteria. The loss of this sugar causes the rigid cell wall to break down, leaving the cell without support. Cells in this situation are vulnerable to external assaults and risk spilling their contents into the extra- cellular space. The organisms evolved two strategies for coping with the loss of muramic acid and thus diverged into two subsequent lines of descent. A new group of prokaryotes, now called archebacteria, developed a new type of rigid cell wall built without muramic acid. Cells that did not develop a new type of wall evolved an internal proteinaceous skeleton; that is, a cytoskeleton. It is at this point, according to Cavalier-Smith, that the modem eukaryote got its start.
In Cavalier-Smith's scheme, the key step in the evolution of eukaryotes is the development of the cytoskeleton. Since the cell's structural integrity is maintained by the cytoskeleton, the cell can afford to have a fluid outer membrane. The increased fluidity of the outer membrane allowed the development of two mechanisms, called endocytosis and exocytosis, which prepared the way for all subsequent steps of eukaryotic evolution.
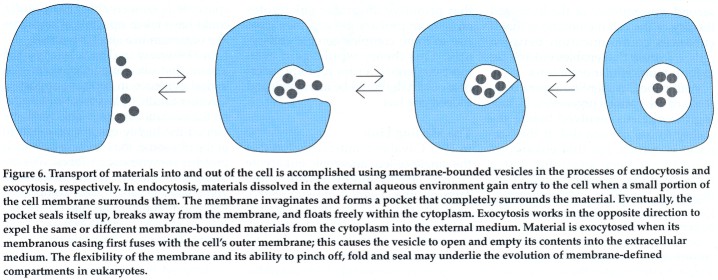 Endocytosis and exocytosis are complementary processes whereby substances can enter (endocytosis) or exit (exocytosis) the cell through membrane-bounded vesicles. In endocytosis, incoming material enters the cell in a vesicle formed by the invagination of a small segment of the outer membrane; the invagination pinches off and seals itself up to form a vesicle, which carries its contents into the cell's interior. In the reverse process of exocytosis, particles leave the cell when the membrane of the vesicle enclosing them fuses with the outer cell membrane. In this case, the vesicle's contents are emptied into the extracellular space. The invaginations of the outer membrane could have given rise to the internal membranous structures of the eukaryotes, such as the cell nucleus, the endoplasmic reticulum and lysosomes. A second consequence of the increased fluidity of the cell membrane is that it enhances communication between cells, a feature that enables eukaryotes to form multicellular organisms.
Endocytosis and exocytosis are complementary processes whereby substances can enter (endocytosis) or exit (exocytosis) the cell through membrane-bounded vesicles. In endocytosis, incoming material enters the cell in a vesicle formed by the invagination of a small segment of the outer membrane; the invagination pinches off and seals itself up to form a vesicle, which carries its contents into the cell's interior. In the reverse process of exocytosis, particles leave the cell when the membrane of the vesicle enclosing them fuses with the outer cell membrane. In this case, the vesicle's contents are emptied into the extracellular space. The invaginations of the outer membrane could have given rise to the internal membranous structures of the eukaryotes, such as the cell nucleus, the endoplasmic reticulum and lysosomes. A second consequence of the increased fluidity of the cell membrane is that it enhances communication between cells, a feature that enables eukaryotes to form multicellular organisms.
Within the Cavalier-Smith paradigm, some of the various organelles, including the nucleus, evolved before the ability to use oxygen did. If this scenario is correct, the first eukaryotes would have been anaerobic. Later, these primitive anaerobes could have ingested a free-living, oxygen-using organism, via endosymbiosis, to give rise to aerobic eukaryotes.
Scientists often consult the fossil record when considering issues of evolution. In this matter, the record would appear to support some of the ideas of Cavalier-Smith. Prokaryotic cells were present on the earth 3.5 to 4 billion years ago, and eukaryotic cells did not appear until 1.4 to 1.5 billion years ago. Eukaryotic cells that contained mitochondria and were oxygen users, however, were not in evidence until as recently as 850 million years ago. Data from the fossil record reinforce the idea that primitive anaerobic eukaryotes arose from prokaryotes and then gave rise to more complex aerobic eukaryotes. This theory would gain even more credence if an appropriate anaerobic eukaryote could be identified, and indeed one has.
The Missing Link
In 1987 Cavalier-Smith suggested that the single-celled, anaerobic eukaryote Giardia lamblia might in fact be the missing link. Giardia, he suggested, might be the anaerobic eukaryote that ingested an oxygen-using bacterium; this union could have yielded the first aerobic eukaryote, from which all others may have descended. He based his prediction on an analysis of the types and structures of the organelles in Giardia. One important observation is that Giardia lacks mitochondria and is an obligate anaerobe. In further support of this notion, experiments in which Giardia ingested a marker molecule visible in electron micrographs show that the organism is capable of endocytosing particles from the extracellular
space. It is conceivable that Giardia could have taken up a mitochondrion-like organism in a similar manner.
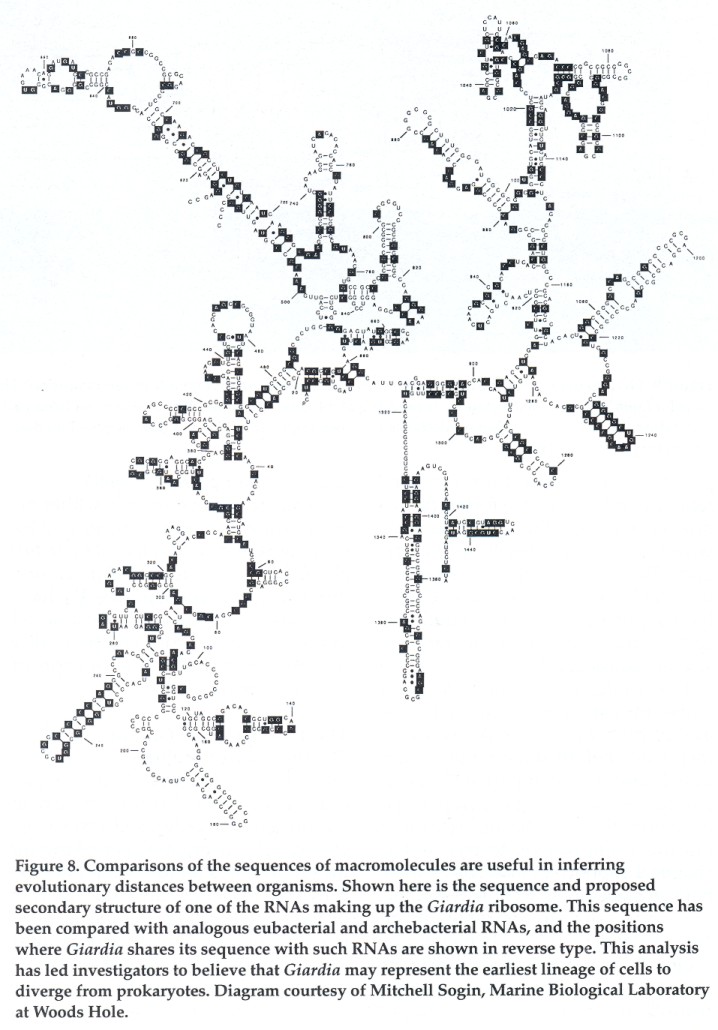 In 1989 one of us (peattie) was part of a team that uncovered molecular-biological evidence in further support of Cavalier-Smith's placement of Giardia on the evolutionary tree. This work exploited the highly conservative nature of the ribosome, the structure on which proteins are synthesized. Ribosomes are found in all known cells-prokaryotic and eukaryotic alike. Their component RNA and protein molecules are assembled and folded to create a characteristic shape. There has been relatively little change over time in the molecules making up the ribosome. As a result, prokaryotic and eukaryotic ribosomes are fairly similar. The conserved nature of these molecules and their ubiquity makes them extremely useful for inferring evolutionary distances between organisms. Comparing the sequence of nucleotides in a ribosomal RNA shows how much the molecule has changed in the course of evolution from one organism to another.
In 1989 one of us (peattie) was part of a team that uncovered molecular-biological evidence in further support of Cavalier-Smith's placement of Giardia on the evolutionary tree. This work exploited the highly conservative nature of the ribosome, the structure on which proteins are synthesized. Ribosomes are found in all known cells-prokaryotic and eukaryotic alike. Their component RNA and protein molecules are assembled and folded to create a characteristic shape. There has been relatively little change over time in the molecules making up the ribosome. As a result, prokaryotic and eukaryotic ribosomes are fairly similar. The conserved nature of these molecules and their ubiquity makes them extremely useful for inferring evolutionary distances between organisms. Comparing the sequence of nucleotides in a ribosomal RNA shows how much the molecule has changed in the course of evolution from one organism to another.
Change is thought to be a function of time. If a great deal of time elapses between the emergence of organisms, there is more opportunity for the sequences of their nucleic acids and proteins to change. If two organisms evolve in a shorter space of time, there are fewer opportunities for the sequences to change. By analyzing the nucleotide sequences of ribosomal RNAs from different organisms and determining the degree of similarity or difference between these RNAs, we can reach some conclusions about how closely or distantly the organisms are related. With the aid of a computer, the sequences can be organized into a phylogenetic tree whose branching order indicates the likely order in which the organisms diverged from some common ancestor. Such phylogenetic trees can help clarify the order of branching among prokaryotes and eukaryotes and assess which organism represents the earliest line of descent among the eukaryotes.
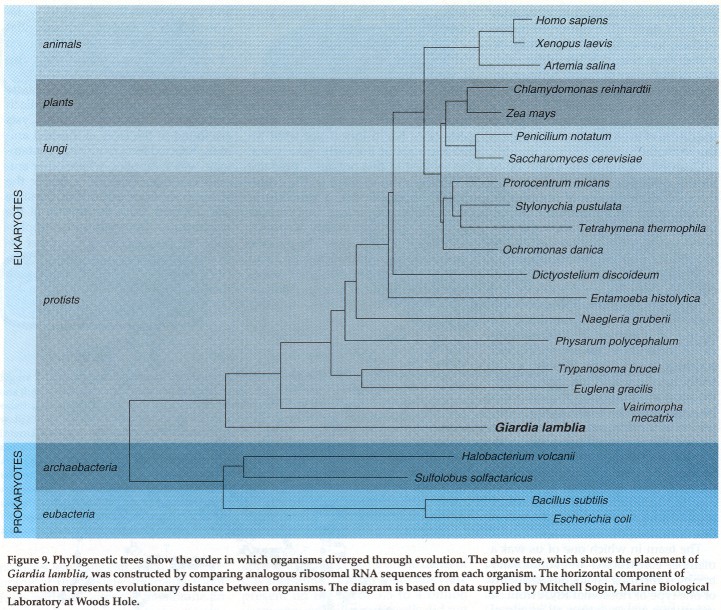 The team in which one of us was a member constructed an evolutionary tree by comparing analogous segments of one type of ribosomal RNA from 54 organisms representing all levels of evolution. By this method we discovered that Giardia lamblia has a ribosomal-RNA sequence that is very close to that of prokaryotic cells. This Giardia ribosomal RNA shares more of its sequence with prokaryotes than does the
corresponding RNA of any other eukaryote. Thus Giardia is evolutionarily closer to the prokaryotes than other eukaryotes are, and we can take this to mean that Giardia is a member of the earliest emerging eukaryotic lineage.
The team in which one of us was a member constructed an evolutionary tree by comparing analogous segments of one type of ribosomal RNA from 54 organisms representing all levels of evolution. By this method we discovered that Giardia lamblia has a ribosomal-RNA sequence that is very close to that of prokaryotic cells. This Giardia ribosomal RNA shares more of its sequence with prokaryotes than does the
corresponding RNA of any other eukaryote. Thus Giardia is evolutionarily closer to the prokaryotes than other eukaryotes are, and we can take this to mean that Giardia is a member of the earliest emerging eukaryotic lineage.
Given that Giardia lacks mitochondria and is necessarily anaerobic, its phylogenetic position provides compelling evidence for Cavalier-Smith's hypothesis that the anaerobic eukaryotes preceded the aerobes. At the same time, this evidence conflicts with Margulis's position that eukaryotes evolved in response to the buildup of oxygen on the primitive earth. The ability to use oxygen would appear to have arisen later, rather than earlier, in the progression from prokaryote to eukaryote.
Valuable Lessons
Having established that Giardia occupies an important position in the transition from prokaryotes to eukaryotes, we have been interested in exploring aspects of its life history to see what we can learn about the early eukaryotes. But interest in this organism predates the discovery of its place on the evolutionary tree. Giardia, a unicellular organism, is a major intestinal parasite capable of infecting a variety of species, including human beings. The parasite can be found in both developed and underdeveloped countries, and causes diarrhea, abdominal cramps, malaise and weight loss. Because of its wide-spread distribution and the potential severity of infections, Giardia has been of interest to parasitologists and epidemiologists for some time.
There are two phases in the Giardia life cycle: trophozoites and cysts. Each cyst contains two trophozoites. Cysts are found in the feces of infected animals. Infected fecal matter can contaminate water supplies, where other animals can ingest the cysts. Inside the stomach, cysts are exposed to digestive acids, which cause the release of the trophozoites. Once released, a trophozoite attaches to cells of the upper intestine by means of a disc on the parasite's ventral surface. The parasites are thought to remain for a time in the upper intestine, where they feed and replicate. When they are forced further down the intestinal tract into the small bowel or colon, the trophozoites form cysts. These cysts are then excreted and the cycle is repeated. Because Giardia has the potential to infect a number of species, the possibilities for transmission are considerable.
The Giardia trophozoite lacks many of the subcellular organelles characteristic of higher aerobic eukaryotes. As mentioned before, it has no mitochondria. It also has no apparent Golgi apparatus. Some investigators report seeing a primitive endoplasmic reticulum, but this claim has yet to receive biochemical support. Giardia has an internal cytoskeleton as well as lysosomes that contain digestive enzymes. Its genetic material is encased in membrane-enclosed compartments. In our laboratory, studies are carried out using these trophozoites, graceful, tear-shaped cells with four pairs of flagella.
Of all the cellular features within the trophozoite, the most puzzling and intriguing is that Giardia has not one nucleus, but two nuclei. Furthermore, these nuclei are equal in size. We have undertaken a study of the dual nuclei in the hope of contributing to a fuller understanding of the evolution of higher eukaryotic cells.
The Two Nuclei of Giardia
The presence of multiple nuclei is not unique to Giardia. Several other single-celled eukaryotes also have more than one nucleus, but only Giardia and a few closely related organisms making up the group called diplomonads have exactly two nuclei of equal size. In light of the evolutionarily significant position Giardia holds, we have been most eager to understand the importance of this unusual nuclear configuration. We have been trying to elucidate the structural and functional contributions made by the two nuclei.
Recently, we have shown that the two nuclei are not only equal in size, but also that they contain the same amount of DNA. In addition, using techniques that enable us to see the nucleic acids DNA and RNA under the microscope, we have demonstrated that each nucleus contains four major chromosomes.
We were then curious to learn whether the DNA in each nucleus encodes the same information. Since we know the sequence of one ribosomal RNA within the cell, and we know that the template for this RNA is archived in the DNA, we probed the DNA in each nucleus to see whether each contained this ribosomal sequence. We found that each nucleus did indeed contain sequences specifying this ribosomal RNA.
The mere presence of the sequence in each nucleus, however, does not necessarily mean that each one is used as a template for ribosomal RNA. Thus we sought to establish whether the ribosomal RNA in the cell could have been derived from the DNA in either nucleus. We tagged all nascent RNAs with a radioactive label, and found radioactive RNA emerging from both nuclei. Since ribosomal RNA is by far the most abundant RNA species, we can infer that it was being copied from DNA templates in both nuclei. This suggests that the DNA in both nuclei is functionally equivalent and equally likely to serve as the template for the ribosomal RNA that makes its way out of the nucleus and into the cytoplasm.
Additional studies conducted on other multinucleated cells suggest that the DNA in these is not functionally equivalent. For example, in two well-studied single-celled organisms, Tetrahymena and Paramecium, there are unequal-size nuclei. Tetrahymena has one large nucleus and one small nucleus. The smaller nucleus contains two copies of the genome and is therefore diploid. The DNA in this micronucleus is not used as a template, but is passed on to progeny cells. The small nucleus then develops into a large nucleus, and only then does its DNA assume a template role. Paramecium has a large nucleus and two small diploid nuclei, which are likewise only passed on to future generations.
Since the arrangement of nuclei in Giardia is anomalous, even among multinucleated cells, the question arises: What is the possible evolutionary significance of the two equal-size nuclei of Giardia and their contents? We have already noted that we detect four major chromosomes in each nucleus, and others have reported that there are between four and five major chromosomes in the organism. The amount of DNA in each cell, and its complexity, have been determined. Taken together, these data and other experiments suggest that each nucleus of Giardia is haploid; that is, each contains a single representation of the organism's genetic information. The entire trophozoite, which contains two nuclei, would therefore be diploid.
The diploid state can be greatly advantageous, and most highly evolved, complex organisms have adopted this arrangement. If a cell has only a single copy of genetic information, any alteration or mutation of that information could result in nonfunctional proteins, with dire and possibly lethal consequences for the cell. But if the cell has two sets of instructions and one of them becomes nonfunctional, the second set can serve as a backup, and may compensate for the loss of the first. Furthermore, if a segment of the first set undergoes a mutation that provides a new beneficial function, the other copy can still perform the original function. With only one set of instructions, the organism risks losing an existing function to gain a new one. Again, this could ultimately lead to the cell's demise. A diploid organism has the advantage of retaining the old while developing new, advantageous functions.
The putative haploidy of each of the Giardia nuclei is intriguing with respect to the organism's evolutionary importance. It is possible that a single haploid nucleus gave rise to a second identical nucleus, thus giving the entire organism the various advantages of the diploid state. Later in evolution, the two haploid nuclei could have fused to produce the sole diploid nucleus characteristic of most higher eukaryotes. This hypothetical scenario explains the transition of a haploid prokaryote to a diploid eukaryote. It also predicts that the higher eukaryotes that contain a single diploid nucleus are the evolutionary descendants of a binucleated eukaryote.
Bibliography
Alberts, B., D. Bray, J. Lewis, M. Raff, K. Roberts and J. D. Watson. 1989. Molecular Biology of the Cell. Second Edition. Garland Publishing, Inc. New York.
Bockman, D. E., and W. B. Wiborn. 1968. Electron microscopic localization of exogenous ferritin within vacuoles of Giardia muris. Journal of Protozoology 15:26-30.
Boothroyd, J. C., A. Wang, D. A. Campbell and C. C. Wang. 1987. An unusually compact ribosomal DNA repeat in the protozoan Giardia lamblia. Nucleic Acids Research 15:4065-4084.
Cavalier-Smith, T. 1975. The origin of nuclei and of eukaryotic cells. Nature 256:463-468.
Cavalier-Smith, T. 1981. The origin and early evolution of the eukaryote cell. Symposia of the Society of General Microbiology 32:33-84.
Cavalier-Smith, T. 1987a. Eukaryotes with no mitochondria. Nature 326:332-333.
Cavalier-Smith, T. 1987b. The origin of eukaryote and archaebacterial cells. Annals of the New York Academy of Sciences 503:17-54.
Cavalier-Smith, T. 1988. Origin of the cell nucleus. BioEssays 9:72-78.
Cavalier-Smith, T. 1989. Archaebacteria and Archezoa. Nature 339:100-101.
Doolittle, W. F. 1988. Bacterial evolution. Canadian Journal of Microbiology. 34:547-551.
Dyer, B. D., and R. Obar, eds. 1985. The Origin of Eukaryotic Cells. Van Nostrand Reinhold Co. New York.
Feely, D. E. 1985 Histochemical localization of acid phosphatase in Giardia. Anatomical Records 202:54A
Friend, D. S. 1966. The fine structure of Giardia muris. The Journal of Cell Biology 29:317-332.
Halvorson, H. 0., and A. Monroy, eds. 1984. The Origin and Evolution of Sex. MBL Lectures in Biology. Volume 7. Alan R. Liss, Inc. New York.
Kabnick, K. S., and D. A. Peattie. 1990. In situ analyses reveal that the two nuclei of Giardia lamblia are equivalent. Journal of Cell Science 95:353-360.
Lindmark, D. G. 1988. Giardia lamblia: Localization of hydrolase activities in lysosome-like organelles of trophozoites. Experimental Parasitology 65:141-147.
Margulis, L., 1981. Symbiosis in Cell Evolution. Life and its environment on the early earth. W. H. Freeman and Co. New York. Margulis, L. and D. Sagan. 1986. Microcosmos. Summit Books, New York.
Nemanic, P. C., R. L. Owen, D. P. Stevens, and J. C. Mueller. 1979. Ultrastructural observations on giardiasis in a mouse model. II Endosymbiosis and organelle distribution in Giardia muris and Giardia lamblia. Journal of Infectious Diseases 140:222-228.
Preer, J. R., Jr. 1989. Update on the molecular genetics of Paramecium. Journal of Protozoology 36:182-184.
Prescott, D. M., and G. E. Stone. 1967. Replication and function in the protozoan nucleus. In Research in Protozoology, vol. 2. (T.- T. Chen, ed.), pp. 117-146. Pergamon Press, New York.
Roberts-Thompson, I. C. 1984. Giardiasis. In Tropical and Geographical Medicine, K. S. Warren and A. A. F. Mahmoud, eds., McGraw-Hill, New York. pp. 319-325.
Sagan, L. 1967. On the origin of mitosing cells. Journal of Theoretical Biology 14:225-274.
Shih, M.-C., P. Heinrich, and H. M. Goodman. 1988. Intron existence predated the divergence of eukaryotes and prokaryotes. Science 242:1164-1166.
Sogin, M. L., J. H. Gunderson, H. J. Elwood, R. A. Alonso and D. A. Peattie. 1989. Phylogenetic meaning of the kingdom concept: An unusual ribosomal RNA from Giardia lamblia. Science 243:75-77.
Sonneborn, T. M.1974a. Tetrahymena pyriformis. In Handbook of Genetics, vol. 2: Plants, Plant Viruses, and Protists (R. C. King, ed.), pp. 433-467. Plenum Press, New York.
Sonneborn, T. M. 1974b. Paramecium aurelia. In Handbook of Genetics, vol. 2: Plants, Plant Viruses, and Protists (R. C. King, ed.), pp. 469-594. Plenum Press, New York.
Weisehahn, G. P., E. J. Jarroll, D. G. Lindmark, E. A. Meyer, and L. M. Hallick. 1984. Giardia lamblia: autoradiographic analysis of nuclear replication. Experimental Parasitology 58:94-100.
Woese, C. R. 1987. Bacterial evolution. Microbiological Reviews 51:221-271.
Woodard, J., E. Kaneshiro, and M. A. Gorovsky. 1972. Cytochemical studies on the problem of micronuclei in Tetrahymena. Genetics 70:251-260.
Yao, M.-C., and M. A. Gorovsky. 1974. Comparison of the sequences of macro- and micronuclear DNA of Tetrahymena pyriformis. Chromosoma 48:1-18.
Karen Kabnick received her Ph.D. in biology from MlT. She currently works with Debra Peattie as a postdoctoral fellow at the Harvard School of Public Health. In addition, she teaches cell biology in the Division of Continuing Education at Harvard University.
Debra Peattie is Adjunct Assistant Professor
in the Department of Tropical Public Health at the Harvard School of Public Health and Staff Scientist at Vertex Pharmaceuticals Incorporated in Cambridge, Massachusetts. She obtained a B.A. in chemistry at Hollins College, Virginia, and a Ph.D. in biochemistry and molecular Biology at Harvard University. She is delighted that Giardia lamblia occupies a significant niche in biological evolution and is very pleased never to have suffered the pangs of giardiasis.