

Circular overlaps: Rare demonstrations of speciation
|
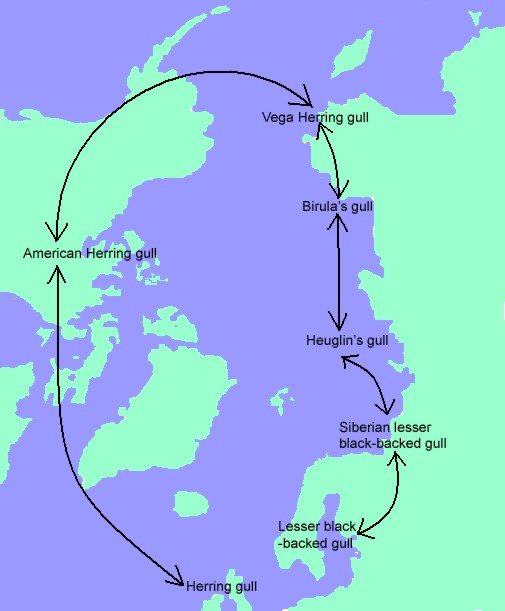
One of the most widely known ring species is the series of taxa in the Herring Gull (Larus argentatus) and Lesser Black-backed Gull (L. fuscus) group. The races of this species are arranged in a circumpolar ring, but L. argentatus, coming from America, invaded western Europe and lives now side by side with L. fuscus, like a good species, although the two "species" are connected by a chain of intermediate forms in Siberia and North America.
Those conclusions were based primarily on morphological variation. The two supposedly terminal forms, which overlap in distribution in Europe, differ in size, shape, and plumage color, but those traits gradually change through the chain of forms encircling the Arctic Ocean.

|
A number of researchers have examined molecular variation from parts of the Larus species complex in the hope of clarifying historical relationships between taxa and current patterns of gene flow. Mitochondrial DNA variation suggest that North American L. smithsonianus is more distantly related to the European forms Herring Gull (L. argentatus argenteus and L. argentatus argentatus) than is the European Lesser Black-backed Gull (L. fuscus). It appears that L. smithsonianus is highly related to a group of several other morphologically divergent species, such as Thayer's Gull (L. thayeri), Glaucous Gull (L. hyperboreus), Iceland Gull (L. glaucoides), Slaty-backed Gull (L. schistisagus), and California Gull (L. californicus), most of which are also found primarily in North America. Researchers suggest that L. smithsonianus, which is usually treated as a subspecies of L. argentatus (e.g. Harrison 1985, Burger and Gochfeld 1996), should be treated as its own species, Larus smithsonianus. This conclusion should be confirmed with higher sample sizes (more than the initial nine smithsonianus samples from two sites), before modifying the theory that European L. argentatus resulted from a circumpolar expansion and gradual divergence of L. fuscus.
THE RING SPECIES CONCEPTS
First, the presence of a series of intermediate forms between two species shows that variation between species is qualitatively similar, though different in degree, as variation within a species. Ring species demonstrate that small changes can eventually accumulate into the differences between distinct species.
Second, ring species may be used to reconstruct the history and causes of divergence during speciation. If an ancestral species expands around both sides of an ecological barrier, the expanding fronts might diverge to the level of species by the time they meet on the other side of the barrier. If a ring species arose in that way, variation around the ring might represent variation in time, allowing the observation of the traits of two species as well as their common ancestor.
Third, ring species potentially show that speciation between the terminal forms can occur despite substantial gene flow around the ring. Such a demonstration would show that speciation does not require geographic isolation, something that has often been considered an essential condition for speciation. Genetic exchange between populations generally inhibits divergence, but recent theoretical and empirical studies have shown that substantial divergence can occur even with gene flow, especially when there is natural selection for local adaptation. There is no theoretical reason why ring species with extensive gene flow through the ring cannot exist.
PROPOSED CASES OF RING SPECIES
At least 23 taxa have been proposed as ring species: birds (12), amphibians (2), rodents (3), arthropods (5), and plants (1). The most ideal example of a ring species would have three characteristics. First, two distinctive forms would coexist while being reproductively isolated in the same area. Second, there would be a chain of intermediate forms connecting them through a continuous geographic ring. Third, there would be no breaks in gene flow except between the terminal forms. But as our discussion of the ring species concept above shows, biologists might consider a system to be a ring species even if it did not have all of those characteristics.
In fact, few if any of the proposed examples have all the characteristics of ideal ring species. Most of the proposed cases have major gaps in distribution, making significant gene flow throughout the chain of populations unlikely. Also, in many proposed ring species there is either hybridization between the supposed terminal forms of the ring or locations around the ring where there is sudden change between forms that are reproductively isolated to some degree.
Nevertheless, all the cases have one characteristic in common: two distinct forms coexist, but forms elsewhere have intermediate characteristics. For example, in the Galapagos Islands, the Medium Tree-finch (Camarhynchus pauper) and the Large Tree-finch (C. psittacula psittacula) coexist on Floreana, but a series of intermediate forms occur on nearby islands (C. psittacula affinis on Isabela, affinis/psittacula intergrades on Duncan, C. psittacula psittacula on Santa Cruz. The progressive morphological differences in those finches through the chain of islands, with the most divergent forms coexisting as separate species on the same island, illustrates how small changes can accumulate into differences between species. This link between microevolution and speciation is particularly clear when the history of range expansion can be reconstructed, in which case we can use variation in space to infer variation in time.
Molecular analyses of microsatellite loci and allozyme variation resulted in consistent phylogenies based on morphological characters. An unexpected result was that genetic divergence among Galápagos Island populations of the warbler finch (Certhidea olivacea) predates the radiation of all other Darwin's finches. This deep split is surprising in view of the relatively weak morphological differentiation among Certhidea populations and supports the hypothesis that the ancestor of all Darwin's finches was phenotypically similar to Certhidea. The results also resolve a biogeographical problem: the Cocos Island finch evolved after the Galápagos finch radiation was under way, supporting the hypothesis that this distant island was colonized from the Galápagos Islands. Monophyletic relationships are supported for both major groups, the ground finches (Geospiza) and the tree finches (Camarhynchus and Cactospiza), although the vegetarian finch (Platyspiza crassirostris) appears to have diverged prior to the separation of ground and tree finches.
|
THE LARUS COMPLEX
The mitochondrial patterns as presently understood do not support the hypothesis that the fuscus-smithsonianus-argentatus circumpolar ring arose by a simple history of gradual expansion and divergence. Morphologically, the gulls still show a remarkable pattern, with stepwise changes through the chain of forms and two clearly distinct forms living together in Europe. If the morphological similarity between European argentatus and North American smithsonianus did not arise as a result of recent shared ancestry, it must have arisen through convergence or from morphological stasis while other related groups speciated and diverged morphologically. The low amount of molecular variation and the lack of a strong link between molecular relationships and morphological patterns in the Larus group suggest that phenotypic evolution can occur quickly in these gulls. The relative importance of natural selection, sexual selection, and drift in driving those changes remains to be seen. Further study of the Larus gulls could reveal much about the interactions between natural selection, sexual selection, hybridization, and reproductive isolation in speciation.
Irwin, Darren E, and Jessica H Irwin. 2002. Circular overlaps: Rare demonstrations of speciation. The Auk 119(3):596-602
Additional Citations:
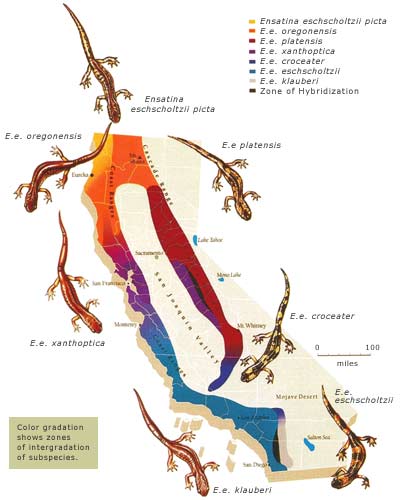
Ensatina eschscholtzi
Ring Species: Salamanders
Genic Variability And Differentiation In The Galapagos Finches (abstract; full article)