Recent developments in molecular biology and new interpretations of the fossil record are gradually altering and adding to the synthetic theory, for 40 years the standard view of the process of evolution.
By G LEDYARD STEBBINS and FRANCISCO J AYALA
To this Darwinian foundation the architects of the synthetic theory, who included the geneticist Theodosius Dobzhansky, the biogeographer and systematist Ernst Mayr, the paleontologist George Gaylord Simpson, the biologist Julian Huxley and one of us (Stebbins), added new elements. The science of genetics made it possible to identify the determinants of the traits on which natural selection acts as genes: heritable units of information governing structure, development and function. Variant traits were held to result from mutations, or lasting alterations that arise at random in individual genes. From population biology came a second new feature of the synthetic theory: an emphasis on the importance of population structure and distribution in the development of new species. The synthetic theory also incorporated the biological concept of species, which leans more heavily on reproductive isolation (an inability to interbreed) than on visible differences in distinguishing species. Although some biologists initially resisted the synthetic theory, for four decades most evolutionists have considered it the best explanation of evolutionary processes, and it has taken a central place in biology.
In the 1970's and 1980's new developments confronted the synthetic theory. An explosion of investigation into the structure of DNA, the carrier of genetic information, has enabled biologists to study the mechanisms of evolution at the molecular level. The new work has thereby amplified the synthetic theory much as the discovery of genes amplified Darwinism.
The molecular studies have also led to two direct challenges to the synthetic theory. One is a proposal that a kind of molecular determinism, rather than pure chance, impels the development of variations in DNA. The other is a contrasting claim, known as the neutral theory, that chance governs not only the initial appearance of genetic variations but also their subsequent establishment in a population. A different kind of challenge, based on new interpretations of the fossil record, has emerged from paleontology. Known as punctuated equilibrium, it holds that evolution proceeds not at a steady pace but irregularly, in fits and starts.
At the outset it must be said that - unlike attacks by creationists and other nonscientists - none of these challenges denies that evolutionary change occurs, that current species have descended from common ancestors or that Darwinian natural selection plays an important part in the process. The disputes are conflicts of degree and emphasis within a shared evolutionary outlook. We believe, moreover, that with modifications both to the traditional views and to the competing theories most of the challenges can be accommodated within the encompassing vision of the synthetic theory.
Molecular Evolution
The most dramatic changes in thinking about evolution stem from new knowledge about genetic processes at a molecular level, and yet many of the implications of that knowledge for evolutionary theory remain obscure. The molecular pathways that lead from genes to visible characteristics are long, complex and as yet largely unexplored. Until the relation of genes to development is better understood at a molecular level the full impact of molecular biology on evolutionary theory cannot be assessed.
Certain consequences of the recent findings are already evident, however. They have shown genetic variation to be far more complex than was thought. involving changes in the number and configuration of genes as well as mutations in individual genes. Mutations and structural changes are now known to affect not only genes but also sequences of DNA that cannot be called genes because they do not code for proteins. Even as the new molecular biology has complicated the traditional picture of genetic variation, it can also be said to have buttressed the synthetic theory by supplying mechanisms underlying many processes the theory invokes. The meaning of evolution at a molecular level is beginning to come clear. It is now possible, for example, to give incipient answers to the question: How do new genes arise?
 The importance of the development of new genes (and not simply the modification of old ones) in the process of evolution is reflected in the relation between the amount of genetic material and the complexity of organisms. The genome, or total genetic complement, of a virus amounts to between 1,300
and 20,000 nucleotide pairs (np). (Nucleotides are the subunits of DNA, which are strung together in pairs to form the two strands of the double helix.) The genome of bacteria includes, on the average, about four million np. Among eukaryotes (organisms with distinct cell nuclei) fungi have between 10 and 20 million np per cell; most animals and plants have several billion np per cell.
The importance of the development of new genes (and not simply the modification of old ones) in the process of evolution is reflected in the relation between the amount of genetic material and the complexity of organisms. The genome, or total genetic complement, of a virus amounts to between 1,300
and 20,000 nucleotide pairs (np). (Nucleotides are the subunits of DNA, which are strung together in pairs to form the two strands of the double helix.) The genome of bacteria includes, on the average, about four million np. Among eukaryotes (organisms with distinct cell nuclei) fungi have between 10 and 20 million np per cell; most animals and plants have several billion np per cell.
The relation between the amount of DNA and the complexity of the organism or its developmental pattern is not precise. A few groups of higher plants, salamanders and some primitive fishes have 1010 np per cell, much more than most mammals. The largest amounts of DNA, some 1012 np per cell, are found in eukaryotes that are relatively simple in structure and development, such as certain species of amoeba and the Psilopsida, primitive relatives of the ferns. Such species can have thousands of copies of some genes as well as long stretches of DNA that do not code for proteins and are not considered to be genes at all.
Indeed, segments of DNA with no known function have been found in surprising numbers. In the human genome a sequence called Alu that is about 300 np long is present in some 300,000 copies, corresponding to 3 percent of the total human DNA; another short segment of about 100 np recurs nearly a million times in the mouse genome. The discovery of these seemingly meaningless repetitions led to the speculation that some molecular evolution is deterministic, proceeding in a particular direction that is independent of chance and natural selection. Such sequences are held to have multiplied not through a series of random events but because a kind of "molecular drive" impelled each sequence to reproduce itself within the genome. Mathematical models have indicated, however, that traditional concepts of evolution suffice to explain the proliferation of such segments.
In spite of the exceptions we have noted and in spite of the fraction of total DNA made up of meaningless sequences, the amount of genetic information carried in each cell does in general increase steadily-from bacteria to molds to higher plants and animals. A typical gene consists of a thousand or more nucleotides arrayed in an order as crucial as the sequence of letters in a sentence. A random sequence of a thousand letters is not likely to make sense; they must be organized in order to convey information. How have meaningful sequences of DNA accumulated in the course of evolution?
One way in which an organism's complement of genes can increase abruptly is through polyploidy: a doubling of the number of chromosomes from one generation to the next, producing offspring that usually are reproductively isolated from the parent generation and in effect constitute a new species [see "Cataclysmic Evolution," by G. Ledyard Stebbins, Jr.; SCIENTIFIC AMERICAN, April, 1951]. Some organisms with very large amounts of DNA, such as Psilopsida, are polyploid. Indeed, about 47 percent of all flowering plants are polyploid, including cultivated plants such as the potato, the strawberry and wheat. Nevertheless, many complex plants and animals are not polyploid, and polyploidy does not explain the origin of genes. It is simply a mechanism by means of which existing genes are multiplied.
New Genes
A process that does create entirely new genes is the tandem multiplication, over evolutionary history, of a short sequence of nucleotides. A tandem multiplication is the repetition of a sequence in adjacent sites along a chromosome. The multiplication can occur as matching chromosomes pair during the process of meiosis, the cell divisions that give rise to gametes, or sex cells. As the chromosomes line up they sometimes exchange segments; if the chromosomes have paired out of alignment, the exchange can yield one chromosome in which a particular sequence is repeated. Because it is carried by a sex cell, the repetition will be preserved in the genome of the next generation.
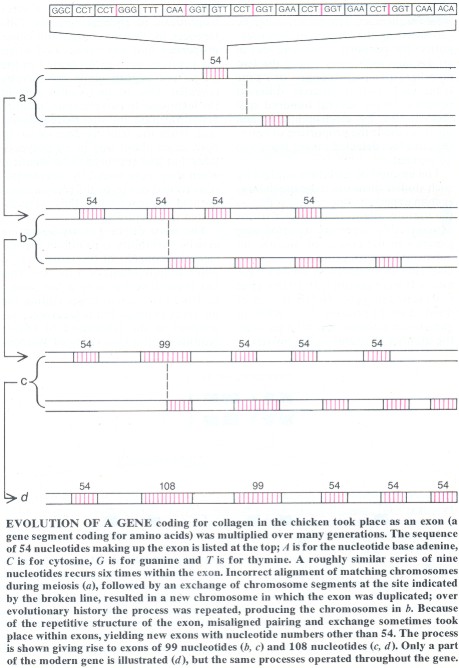 In some cases, such as the a2(I) collagen gene in the chicken, the origin of a gene through tandem multiplication is evident in its structure. Collagen is the main structural protein of bone, cartilage, connective tissue and skin in vertebrates. In chickens the a2(I) gene consists of more than 50 exons, which are discrete sequences of DNA that code for proteins and are separated by noncoding intervening sequences, or introns; the gene has a total length of about 38,000 np. The exons consist of a repeated sequence nine nucleotides long. The repetitions differ somewhat in their component nucleotides, but their common origin can be recognized because the triplet of amino acids for which each sequence codes invariably begins with the amino acid glycine; proline often occupies either or both of the two subsequent positions in the triplet.
In some cases, such as the a2(I) collagen gene in the chicken, the origin of a gene through tandem multiplication is evident in its structure. Collagen is the main structural protein of bone, cartilage, connective tissue and skin in vertebrates. In chickens the a2(I) gene consists of more than 50 exons, which are discrete sequences of DNA that code for proteins and are separated by noncoding intervening sequences, or introns; the gene has a total length of about 38,000 np. The exons consist of a repeated sequence nine nucleotides long. The repetitions differ somewhat in their component nucleotides, but their common origin can be recognized because the triplet of amino acids for which each sequence codes invariably begins with the amino acid glycine; proline often occupies either or both of the two subsequent positions in the triplet.
From the gene's structure its evolutionary history can be reconstructed. A series of five tandem repetitions of the ancestral. 9-np sequence yielded an exon 54 np long. Next the basic exon was multiplied about 50 times. The repetitions have yielded some exons with nucleotide numbers other than 54, but they are always a multiple of nine. Because of the recurrence of the 9-np sequence within each exon, misaligned pairing of chromosomes during meiosis sometimes left corresponding exons overlapping by some multiple of nine. Changed nucleotide numbers resulted when the chromosomes exchanged segments.
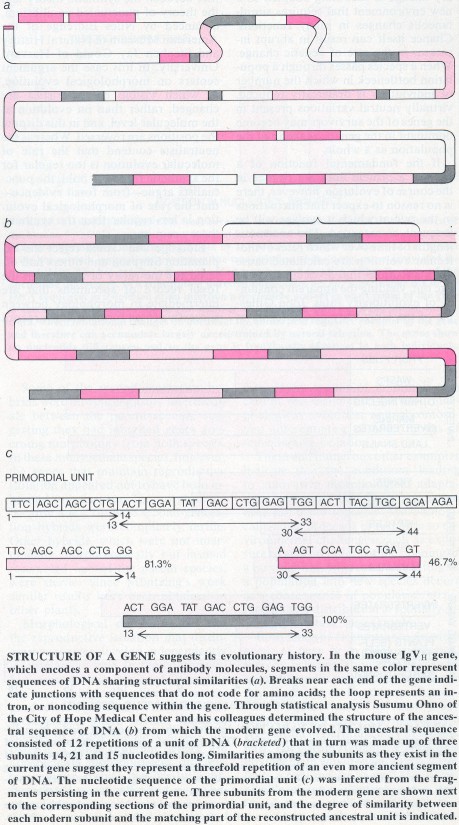 Susumu Ohno of the City of Hope Medical Center and his collaborators have traced a similar origin for some of the genes coding for immunoglobulins, or antibody molecules, in mice. Immunoglobulins are proteins consisting of two large ("heavy") and two smaller ("light") polypeptide chains. In each heavy and light chain a region whose nucleotide sequence is nearly invariant is joined to an extremely variable region; possible combinations of the variable regions yield the enormous diversity of antibodies that serve to protect mammals from foreign substances.
Susumu Ohno of the City of Hope Medical Center and his collaborators have traced a similar origin for some of the genes coding for immunoglobulins, or antibody molecules, in mice. Immunoglobulins are proteins consisting of two large ("heavy") and two smaller ("light") polypeptide chains. In each heavy and light chain a region whose nucleotide sequence is nearly invariant is joined to an extremely variable region; possible combinations of the variable regions yield the enormous diversity of antibodies that serve to protect mammals from foreign substances.
Ohno and his colleagues have found that the genes, each about 600 np long, coding for the variable region of the immunoglobin heavy chain evolved from an ancestral gene that in turn was the result of 12 tandem repetitions of a primordial sequence of 48 np. Further analysis showed that the 48-np building block represents an association of three segments, 14,21 and 15 np long. Although they have been modified by point mutations over the course of evolutionary history, the three segments retain enough similarities to suggest that they represent a tandem triplication of an even smaller building block.
The gene encoding the constant region of an immunoglobulin heavy chain in mice evinces a different evolutionary history, one that may also be common. The heavy chain's constant region includes three protein domains with distinct functions. One domain interacts with cell surfaces, another activates complement (a group of proteins that destroy foreign cells) and a third forms the attachment point for the light chain. In addition the heavy chain has a fourth segment, a hinge region that separates two portions of the molecule. Each of the four exons in the gene codes for one of the four components of the chain, suggesting each represents a small primordial gene that once coded for a separate polypeptide with a function ancestral to that of the modern protein component.
Gene Duplication
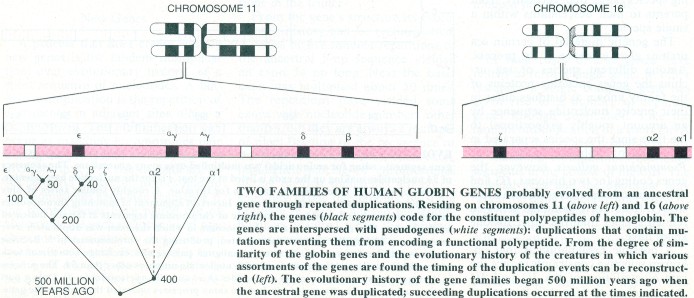 In such cases diverse genes have united to form a single gene. In other instances a single complex gene has given rise to a number of separate genes through duplication. Like the tandem multiplication of a short DNA sequence, the process probably occurs as matching chromosomes exchange segments in the course of meiosis. The original gene and the duplicate may retain the same function; alternatively, one of the genes may preserve the original function while the other evolves a different, albeit related, function. Even before recombinant-DNA techniques were introduced the origin of related genes through the duplication in toto of an ancestral gene was well known; the amino acid sequences of groups of related proteins gave incontrovertible evidence of the common ancestry of the genes coding for them. The globin genes, which code for the four polypeptides that make up human hemoglobin molecules, are an example of a
group of genes for which such an origin is apparent.
In such cases diverse genes have united to form a single gene. In other instances a single complex gene has given rise to a number of separate genes through duplication. Like the tandem multiplication of a short DNA sequence, the process probably occurs as matching chromosomes exchange segments in the course of meiosis. The original gene and the duplicate may retain the same function; alternatively, one of the genes may preserve the original function while the other evolves a different, albeit related, function. Even before recombinant-DNA techniques were introduced the origin of related genes through the duplication in toto of an ancestral gene was well known; the amino acid sequences of groups of related proteins gave incontrovertible evidence of the common ancestry of the genes coding for them. The globin genes, which code for the four polypeptides that make up human hemoglobin molecules, are an example of a
group of genes for which such an origin is apparent.
The globin genes have maintained varying degrees of similarity since the duplication events that gave rise to them. In another evolutionary pattern, a duplicated gene may diverge freely from the original gene because it is superfluous and therefore not subject to the constraints of natural selection. Examples came to light when the length of DNA containing the globin genes was sequenced. Known as pseudogenes, they are sequences homologous to functional genes, from which they arose through duplication, but they contain mutations that prevent them from making a functional polypeptide. The mutations were able to accumulate unhindered by selective pressures because the organism retained a duplicate gene capable of performing the original function. Pseudogenes are now thought to be common in vertebrates and perhaps in other organisms as well.
When new genes arise through duplication, the original and the duplicate gene are usually transmitted together to the descendants of the organism in which the duplication occurred. An apparent twist in the process has been discovered, however-another of the seemingly limitless ways in which evolution proceeds at the genetic level. Occasionally the original gene is found in one species and the duplicate in a totally unrelated organism. The phenomenon is called horizontal DNA transfer, because the DNA is passed from one species to another coexisting species rather than vertically, from parents to their descendants within a single species.
The genetic material of certain sea urchins gives evidence of the process. Among different species of sea urchins the genes encoding proteins of the family known as histones differ in their precise nucleotide sequence by an amount roughly proportional to the time since the species separated in the course of evolution. In the species Psammechinus miliaris, however, the genes coding for two histones, H3 and H4, seem to have diverged much more slowly from corresponding genes in related species than the other histone genes have.
A possible explanation is that unknown selective constraints reduce the rate of evolution of the H3 and H4 genes in Psammechinus to a traction of their rate in other species. An alternative hypothesis is that a cluster of genes coding for H3 and H4 was transferred to Psammechinus from another species, Strongylocentrotus drobachensis. less than a million years ago; the original and the duplicate gene cluster have evolved since then at the usual rate. Because the last common ancestor of the two urchin species lived some 65 million years ago, the histone genes not affected by gene transfer have diverged much further.
The actual mechanisms of horizontal gene transfer are not known. Perhaps the agents are the small circular bits of DNA known as plasmids, which can carry genetic material from one cell to another. Whatever its mechanism, horizontal gene transfer cannot be common. The genes of an organism are coadapted: new or altered genes are favored by natural selection not only because of the function they perform in isolation but also because they complement other genes. Genes are like the players in an orchestra: however virtuosic they may be as soloists, they must also play the same piece or the effect will be cacophony. The coadaptation of the genome reduces the incorporation of functional foreign genes, if it occurs at all, to the realm of rare evolutionary events.
Variation
Molecular biology has yielded insights into not only genetic change over time but also the precondition for such evolution: genetic variation. Evolutionists have shown that among populations in identical or similar environments, the rate of evolution is proportional to the amount of genetic variation within each population [see "The Mechanisms of Evolution," by Francisco J. Ayala; SCIENTIFIC AMERICAN, September, 1978]. The techniques of molecular biology allow the genetic variation within a population to be gauged much more readily than was possible when the synthetic theory was formulated.
 Significant measures of the genetic variation within populations became available in the late 1960's. Gel electrophoresis (in which proteins are embedded in a gel and their mobility in an electric field is compared) and other simple techniques made it possible to determine how many forms of a protein exist in a given species, and in what frequencies. The variation can be expressed as the degree of heterozygosity: the proportion of gene loci in an average individual where the two members of a pair of genes, one gene from each parent, encode different proteins. In the several hundred species studied by electrophoresis and other methods the proportion of heterozygous loci detected ranges from 5 to 20 percent.
Significant measures of the genetic variation within populations became available in the late 1960's. Gel electrophoresis (in which proteins are embedded in a gel and their mobility in an electric field is compared) and other simple techniques made it possible to determine how many forms of a protein exist in a given species, and in what frequencies. The variation can be expressed as the degree of heterozygosity: the proportion of gene loci in an average individual where the two members of a pair of genes, one gene from each parent, encode different proteins. In the several hundred species studied by electrophoresis and other methods the proportion of heterozygous loci detected ranges from 5 to 20 percent.
The amount of variation implied by such studies turns out to be much larger than expected. One reason is that as the various alleles (the distinct forms of a variable gene) are assorted into gametes in the course of meiosis, an individual heterozygous at n loci can give rise to 2n different kinds of gametes. If an organism with 10,000 gene pairs is heterozygous for 10 percent of them, it could produce 21,000 different gametes, a number much larger than the number of atoms in the universe.
Moreover, the amount of genetic variation in a population is usually larger than the results of electrophoresis suggest. Because the technique relies on the differential mobility of proteins in an electric field, differences in the amino acid sequence of a protein that do not change its net electric charge may not be detected. Furthermore, not all nucleotide differences in the DNA sequence result in different protein sequences. The reasons are that different triplets of nucleotides can code for the same amino acid and that some nucleotide sequences do not code for amino acids at all.
One way to detect some of the genetic variation thus hidden from ordinary electrophoresis is to digest each polypeptide chain of a protein into its component peptides (sequences of a few amino acids) and then compare the individual peptides by means of electrophoresis or chromatography. Differences in electric charge and other properties that were concealed when the pep tides were incorporated in the longer polypeptide chain are thereby revealed. One of us (Ayala) applied the technique to two enzymes, alcohol dehydrogenase and superoxide dismutase, from the fruit fly Drosophila. In both cases about one in 10 polypeptides that had appeared to be identical when they were tested by electrophoresis turned out to be different in amino acid sequence when they were digested into peptides.
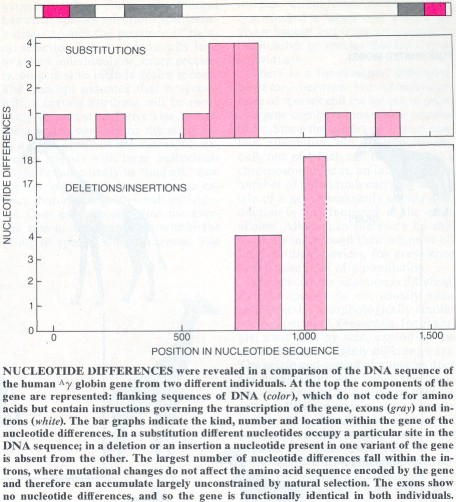 The most direct way to determine genetic variability is to obtain the nucleotide sequence of the same gene in different individuals of a species. This has been done for only a few genes. In 1980 two human genes coding for the same hemoglobin protein were sequenced in the laboratory of Oliver Smithies at the University of Wisconsin at Madison. Although the genes encoded the same product, they differed in their nucleotide sequences by .8 percent if only actual substitutions of one nucleotide for another were counted and by 2.4 percent if nucleotides present in one gene but not in the other were also included. Other genes that have been sequenced, from human beings, mice and fruit flies, have shown comparable degrees of variation. Such results suggest that at the level of the DNA sequence, organisms may be heterozygous at almost all gene loci.
The most direct way to determine genetic variability is to obtain the nucleotide sequence of the same gene in different individuals of a species. This has been done for only a few genes. In 1980 two human genes coding for the same hemoglobin protein were sequenced in the laboratory of Oliver Smithies at the University of Wisconsin at Madison. Although the genes encoded the same product, they differed in their nucleotide sequences by .8 percent if only actual substitutions of one nucleotide for another were counted and by 2.4 percent if nucleotides present in one gene but not in the other were also included. Other genes that have been sequenced, from human beings, mice and fruit flies, have shown comparable degrees of variation. Such results suggest that at the level of the DNA sequence, organisms may be heterozygous at almost all gene loci.
The Neutral Theory
The extensive variation such studies have revealed is one of the bases of the neutral theory, the second of the challenges to the synthetic theory that have emerged from molecular biology. Its chief exponent is Motoo Kimura of the National Institute of Genetics in Japan [see "The Neutral Theory of Molecular Evolution," by Motoo Kimura; SCIENTIFIC AMERICAN, November, 1979]. Neutralists contend that if most genetic differences are under the control of natural selection{as the synthetic theory implies), the observed degree of variation should be low because adaptively beneficial differences should spread through the population and harmful ones should be eliminated. The finding of considerable variation suggests to neutralists that most of the genetic differences neither foster nor hinder an organism's survival and that their persistence or elimination within a population is a matter of chance.
Yet the magnitude of the variation also makes it possible to reconcile the assertion that most differences among alleles are adaptively neutral with the synthetic view, which affirms the importance of Darwinian natural selection. There is so much variation within each generation that even if the majority is neutral, the differences that do have an adaptive effect would supply abundant raw material for the creative force of natural selection.
The question, then, is not which is correct, neutrality or Darwinian natural selection. It is instead how much of the genetic variation persists through chance and how much persists because it confers an adaptive advantage. For examp1e, a mutation of the third nucleotide in a codon (the triplet of nucleotides coding for a particular amino acid) often produces a new codon that is synonymous in translation: both the original and the altered triplet code for the same amino acid. Such "silent" mutations, which do not affect the protein encoded by the gene, might indeed be adaptively neutral, and their frequencies in populations may be due largely to chance. On the other hand, the frequency of the mutation causing the single amino acid difference between the hemoglobin of sickle-cell anemia and normal hemoglobin is clearly under the control of natural selection. When an individual has two copies of the sickle-cell allele, the mutation is fatal, but heterozygosity (possession of one mutant and one normal allele) allows the carrier to be reasonably healthy and confers an adaptive advantage: resistance to malaria.
The Molecular Clock
The neutral theory is based not only on the amount of variation prevailing within a given generation but also on discoveries about the rate of genetic change across generations. Studies relating evolutionary history to the number of molecular differences in DNA sequences or proteins common to several species suggest that a given gene or protein can be regarded as a molecular clock. Its rate of evolution is fairly constant over long periods, and it evolves at much the same rate in different species.
This apparent constancy, the neutralists argue, is incompatible with the notion that molecular change reflects the activity of natural selection. They maintain that the synthetic theory implies variable rates of molecular evolution because selective pressures should vary in intensity over time and from species to species. The chance incorporation of variations into the gene pool of a species is a better way to account for the molecular clock, say the neutralists, because the process would take place at a more or less constant rate.
The synthetic theory does not, however, require the rate of molecular evolution to be as irregular as the critics assume. As long as the function of a gene or a protein is the same in different evolutionary lineages it is not surprising that over millions of years it would appear to evolve at the same rate, since the functional constraints to which it is subject will be much the same. Histone proteins evolve very slowly because they act as a structural support for DNA, and extensive amino acid changes would impair their function; fibrinopeptides, substances involved in blood clotting, evolve much faster because they are not subject to such tight constraints and because the clotting process is often modified in response to environmental changes. The constraints on the evolution of most molecules are intermediate between these two extremes.
The constancy over time of rates of molecular evolution can also be reconciled with the synthetic theory. The number of nucleotide or amino acid substitutions may indeed be greater during a given interval as a result of natural selection, when an evolutionary lineage is becoming adapted to a new environment that requires simultaneous changes in many functions. Chance itself can result in abrupt increases in the rate of genetic change. When a species passes through a population bottleneck in which the number of individuals is drastically reduced, virtually neutral variations present in the genes of the survivors may become dominant in the genetic makeup of the population as a whole.
If the fundamental function of a gene or a protein does not change in the course of evolution, however, there is no reason to expect that fluctuations in the rate at which it evolves will be frequent or extended. The enormous lengths of time over which rates of molecular evolution are calculated cause the fluctuations that do occur to average out, yielding the apparent constancy of evolutionary rates. John Gillespie of the University of California at Davis has constructed mathematical models showing the existence of a molecular clock is compatible with the assumption that molecular evolution is driven by natural selection.
Punctuated Equilibrium
A different kind of question about evolutionary rate is central to the conflict between the synthetic theory and the theory of punctuated equilibrium, advanced by Niles Eldredge of the American Museum of Natural History and Stephen Jay Gould of Harvard University. In this case the argument centers on morphological evolution, in which visible characteristics are changed, rather than on evolution at the molecular level, and in this dispute , the positions are reversed. Whereas the neutralists contend that the rate of molecular evolution is too regular for the synthetic theory to hold, the punctualists argue - from fossil evidence - that the rate of morphological evolution is less regular than the synthetic theory requires.
Eldredge and Gould reject the explanation Simpson and others had offered for the rarity or absence in the fossil record of specimens that are intermediate in morphology between successive fossil forms: that the record is incomplete. Instead they take the record at face value and maintain that the abrupt appearance of new fossil species reflects their development in bursts of evolution, after which the species may have changed little over millions of years. In their view the notion of gradualism must be replaced by a picture in which spurts of change alternate with long periods of stasis.
The dispute with the punctualists loses some pf its focus when one recognizes that it is partly an artifact of a radical difference in time scales: the time scale of the paleontologists who propose the theory of punctuated equilibrium and that of the geneticists who were instrumental in formulating the synthetic theory. Since successive layers in geologic strata may have been laid down tens of thousands of years apart, morphological changes that developed over thousands of generations may make an abrupt appearance in the fossil record. In contrast, geneticists refer to changes that require 200 generations or more as gradual, since they exceed the time span of all experiments except those on microorganisms. In speaking on the one hand of sudden change and on the other of gradual evolution, the punctualists and the gradualists are in many cases talking about the same thing.
 The apparent episodes of abrupt change in the fossil record, then, do not necessarily weigh against the synthetic theory and its emphasis on gradual processes. Can the same be said for the long periods of stasis the punctualists point to, in which the visible characteristics of a species change either little or not at all? Taking a closer look at the fossil record, many paleontologists outside the punctualist camp agree that the stability of visible characteristics over millions of years is much greater than the architects of the mid-century synthetic theory thought at the time. Some of the apparent constancy in the record, however, may reflect a phenomenon known as mosaic evolution, first described by the British evolutionist Sir Gavin de Beer.
The apparent episodes of abrupt change in the fossil record, then, do not necessarily weigh against the synthetic theory and its emphasis on gradual processes. Can the same be said for the long periods of stasis the punctualists point to, in which the visible characteristics of a species change either little or not at all? Taking a closer look at the fossil record, many paleontologists outside the punctualist camp agree that the stability of visible characteristics over millions of years is much greater than the architects of the mid-century synthetic theory thought at the time. Some of the apparent constancy in the record, however, may reflect a phenomenon known as mosaic evolution, first described by the British evolutionist Sir Gavin de Beer.
The term refers to the fact that different parts of an organism do not change at a uniform rate in the course of evolution. Just as genes and proteins do not evolve in lockstep, so the visible characteristics they govern do not evolve at the same rate either. A fossil that lies halfway along a temporal sequence from one organism to the next is not intermediate between the two with respect to every trait. Instead the creature can be compared to a mosaic, resembling the ancestral organism in some characteristics but the descendant in others. Archeopteryx, the fossil species intermediate between the reptiles and the birds, has a skeleton considered to be fully reptilian, but its feathers are birdlike, even to the differentiation between fluffy feathers on the body and longer, stiffer wing feathers. The australopithecines (creatures of two to four million years ago that are ancestral or very nearly ancestral to human beings) resemble humans in that their hipbones are shaped for walking erect, but their cranial capacity is more simian than human.
Some of the stasis apparent in the fossil record may be a product of this evolutionary pattern. When the evolution of a single salient trait is followed in the fossil record, long periods of little change may be evident. While one trait is becalmed by evolution, however, other characteristics that may not be recorded in the fossils can continue to evolve. The punctualist argument may be more convincing with respect to single traits than it is with respect to entire organisms.
Morphology and Speciation
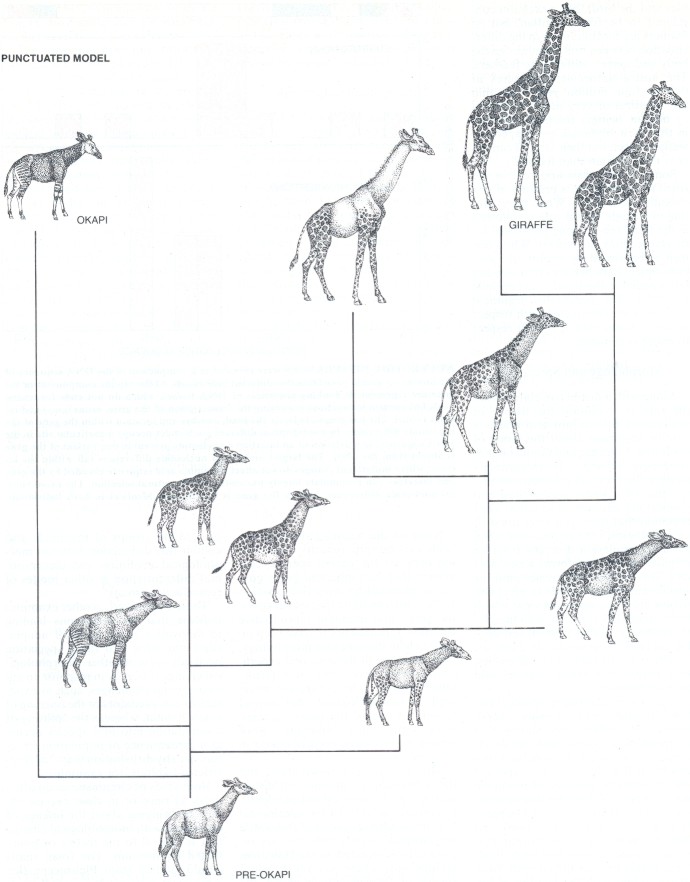 Along with a distinctive claim about the rate of evolution the theory of punctuated equilibrium also makes a specific argument about the mode of evolution. Eldredge and Gould believe anagenesis (changes in morphology along a lineage) and cladogenesis (the splitting of a single species into two species) are causally associated. They maintain that there is a brief burst of morphological change precisely when
a small population diverges from an original species 'and forms a new species; little change then ensues until the species either goes extinct or gives rise to new ones. The opposing view, which the punctualists associate with the synthetic theory, is that gradual morphological change occurs within a species, dividing it into races and subspecies, long before new species can be said to have formed.
Along with a distinctive claim about the rate of evolution the theory of punctuated equilibrium also makes a specific argument about the mode of evolution. Eldredge and Gould believe anagenesis (changes in morphology along a lineage) and cladogenesis (the splitting of a single species into two species) are causally associated. They maintain that there is a brief burst of morphological change precisely when
a small population diverges from an original species 'and forms a new species; little change then ensues until the species either goes extinct or gives rise to new ones. The opposing view, which the punctualists associate with the synthetic theory, is that gradual morphological change occurs within a species, dividing it into races and subspecies, long before new species can be said to have formed.
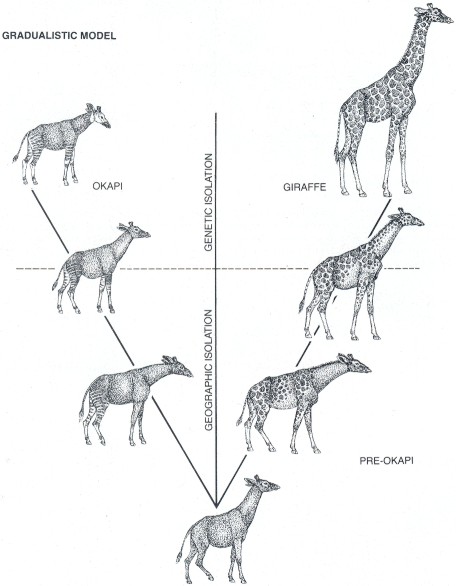 Actually evolutionary change follows both patterns (and many others). Numerous studies have shown that
morphological change and the development of reproductive isolation (the condition that defines a new species) are genetically distinct phenomena: they can occur either together or separately. Arne M�ntzing of the University of Lund showed many years ago that the genes coding for morphological differences among species of flowering plants are not linked with those that interfere with the fertility of hybrids and thereby keep species reproductively distinct. He crossed two species of hemp nettle. The resulting hybrids were partially fertile, enabling Muntzing to produce a second generation of hybrids.
Actually evolutionary change follows both patterns (and many others). Numerous studies have shown that
morphological change and the development of reproductive isolation (the condition that defines a new species) are genetically distinct phenomena: they can occur either together or separately. Arne M�ntzing of the University of Lund showed many years ago that the genes coding for morphological differences among species of flowering plants are not linked with those that interfere with the fertility of hybrids and thereby keep species reproductively distinct. He crossed two species of hemp nettle. The resulting hybrids were partially fertile, enabling Muntzing to produce a second generation of hybrids.
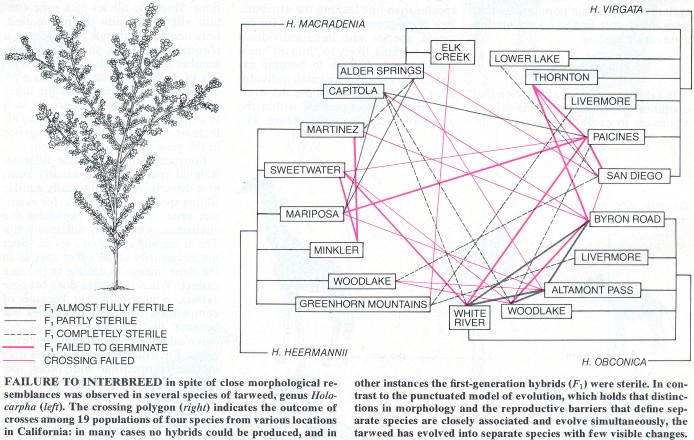 Some of the second-generation hybrids were morphologically intermediate between the parent species, suggesting they had inherited genes governing morphology from both species. In these intermediate species, however, the genes that maintain reproductive isolation appeared not to have been inherited with the genes for morphology, because a few of these second-generation hybrids were completely fertile. Other hybrids, which were not intermediate morphologically but instead resembled one of the parent species, were sterile. Since Muntzing's work similar results have been obtained in other plants.
Some of the second-generation hybrids were morphologically intermediate between the parent species, suggesting they had inherited genes governing morphology from both species. In these intermediate species, however, the genes that maintain reproductive isolation appeared not to have been inherited with the genes for morphology, because a few of these second-generation hybrids were completely fertile. Other hybrids, which were not intermediate morphologically but instead resembled one of the parent species, were sterile. Since Muntzing's work similar results have been obtained in other plants.
Morphological differentiation and the reproductive isolation that distinguishes species develop independently in animals as well. In the continental u.S. many sibling species of Drosophila are morphologically alike but are reproductively-iso1ated. In the Hawaiian archipelago, where several hundred species of Drosophila have arisen, the pattern is quite different. There are conspicuous morphological differences between species that are little differentiated genetically and even between geographically separated, interfertile populations of the same species. Many groups of mammals also exhibit this disjunction between morphological evolution and chromosomal differentiation or other modes of reproductive isolation.
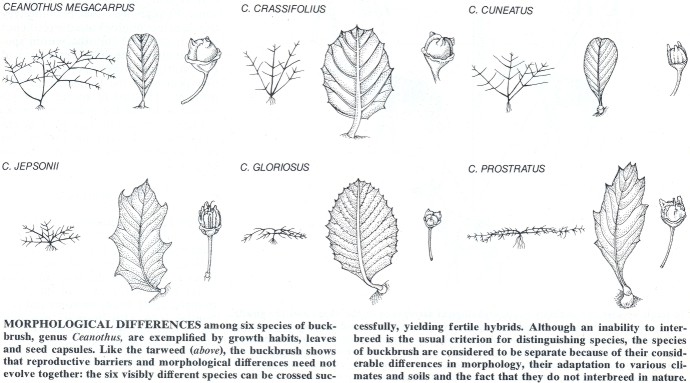 These and numerous other examples indicate that the conditions leading to innovative morphological adaptations and those that favor speciation need not occur together. Morphological change proceeds in response to environmental challenges, such as exposure to new predators or the opening of a new habitat, whereas the splitting of a population into new species occurs as a consequence of population structure: patchy distribution in spatially restricted habitats, for example.
These and numerous other examples indicate that the conditions leading to innovative morphological adaptations and those that favor speciation need not occur together. Morphological change proceeds in response to environmental challenges, such as exposure to new predators or the opening of a new habitat, whereas the splitting of a population into new species occurs as a consequence of population structure: patchy distribution in spatially restricted habitats, for example.
Both kinds of circumstance do often occur at once or in close succession, thereby bringing about the linkage of speciation with morphological change that is central to the theory of punctuated equilibrium. The fossil snails Gould studied from Pleistocene (Ice Age) strata in Bermuda may fit the pattern. They seem to define a branching phylogeny in which each branch represents a new species as well as a recognizable change, although not a profound one, in shell morphology. One must recognize the possibility of circular reasoning, however. A new species is identified each time morphological change is apparent along a temporal sequence of fossils. Hence speciation and morphological evolution are necessarily associated. Actually speciation may have taken place, unrecorded in the fossils, between the recognizable changes, and some of the visible differentiation may define populations that were interfertile and so do not qualify as separate species.
Species Selection
In keeping with their contention that evolution within a species is largely irrelevant to evolutionary trends, the punctualists have embraced a concept known as species selection. Species selection expands the province of natural selection (which is ordinarily held to act on individuals or, more precisely, on genes) to include entire species. The concept assumes that a species with a certain attribute will be more likely to persist and give rise to new species than one lacking the attribute. In a given group of organisms, for example, species with large individuals might be more likely to "bud off" new species, or less likely to become extinct, than species with small individuals. Over evolutionary time the average size of the organisms within the group of species would increase. The increase would result not from evolution toward a larger size within any given lineage but from an increase in the number of species that have large, individuals.
There is a fundamental difference, however, between the comparative fates of species and the spread of genes and gene combinations within populations. Since the alleles of a gene contain alternative bits of information, Only one of which can be present at a chromosomal locus, an increase in the number of individuals carrying one allele of a gene necessarily entails a reduction in the frequency of the other alleles. Alleles can therefore be said to compete, through their adaptive effects on their carriers, for prevalence in the gene pool of a population.
For species the situation is different. Related species do not usually compete directly. Morphologically similar sibling species of Drosophila, for example, exist side by side, exploiting the environment in slightly different ways. The origin and spread of a species does not necessarily cause other species in the same lineage to decline or become extinct. When a species does become extinct, it is usually not because of competition with related species but because its gene pool fails to become adapted to a changing environment. Environmental challenges may include competition with other species, to be sure, but those species are often entirely unrelated to the one that becomes extinct. Insofar as it implies a parallel to the natural selection of genes, then, the notion of species selection is of dubious worth.
It is true, as Simpson recognized some 40 years ago, that evolutionary trends in morphology are the result not only of genetic evolution within species but also of the differential survival and multiplication of species. The former process is the fundamental one, however. The fate of species depends on the ability of the individuals making up the species to cope with the environment, and such an ability can only result from the natural selection of genes.
A New Synthesis
How has the mid-century synthesis weathered recent developments in research and theory? The new molecular biology, by showing that the evolutionary process at the level of DNA is far more complex than had been thought, casts doubt on some old certainties. It has also, however, provided the beginnings of answers to a fundamental matter the architects of the synthetic theory could not address: how genetic information accumulates over evolutionary history. Neutralists and selectionists, strange bedfellows at first glance, can both retain their basic postulates within the harmony of a more comprehensive theory, one that allows chance a greater role in genetic change. Some of the tenets of punctuated equilibrium can be refuted; others are compatible with a modified synthetic theory that encompasses the notions of species stasis and mosaic evolution.
Whatever new consensus emerges from ongoing research and controversy, it is not likely to require rejection of the basic tenets of Darwinism and the mid-century theory. The synthetic theory of the 21st century will differ considerably from the one developed a few decades ago, but the process by which it emerges will be one of evolution rather than upheaval.